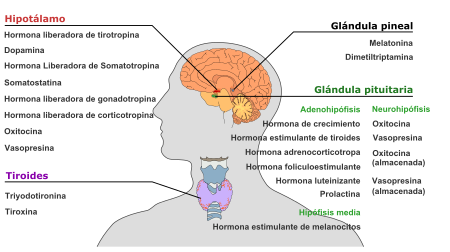
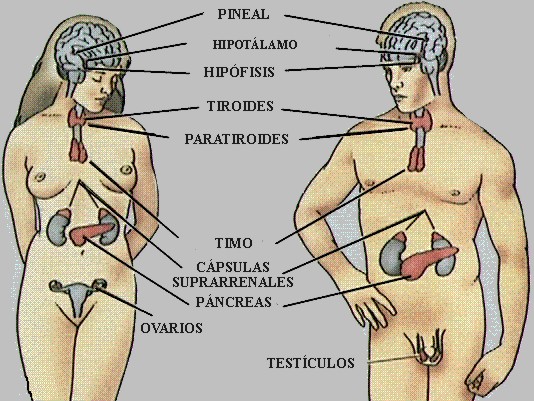
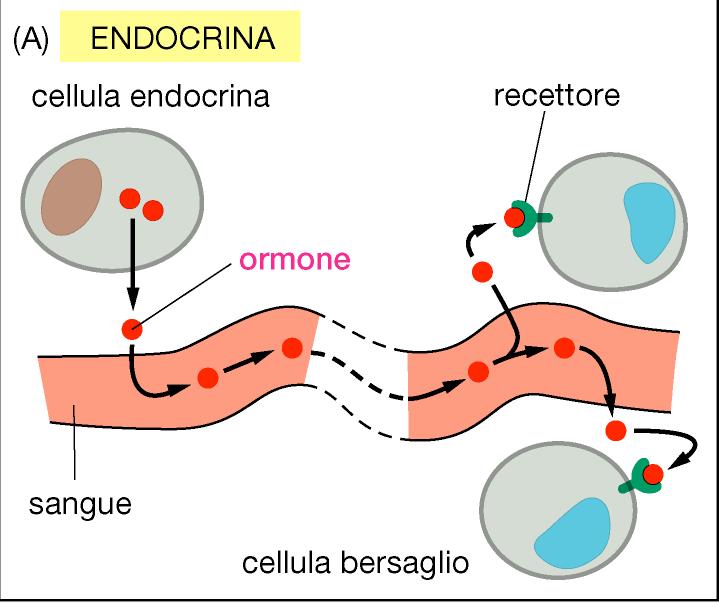
GLÁNDULAS ENDOCRINAS.
Las glándulas endocrinas multicelulares también se pueden
clasificar a partir
de los mismos criterios que se clasifican las glándulas
exocrinas, excepto por el
número de conductos, por carecer de estos. Por consiguiente
se diferencian
por la forma de la porción secretora, la naturaleza de la
secreción y el modo de
segregar.
POR LA FORMA DE LA PORCIÓN SECRETORA.
Las células endocrinas se disponen en forma de cordones,
acúmulos y
folículos en íntima relación con los capilares sanguíneos o
linfáticos hacia
donde vierten el producto de su secreción. En una misma
glándula pueden
presentarse zonas de células con diferente disposición.
En la hipófisis se observan acúmulos celulares, en los
tiroides folículos y en las
glándulas suprarrenales, cordones y acúmulos, por citar
algunos ejemplos.
Fig. 51. Organización histológica de las glándulas
endocrinas en acúmulos, folículos y
cordones.
POR LA NATURALEZA DE LA SECRECIÓN.
Las diferentes hormonas segregadas por las glándulas
endocrinas pueden ser,
por su naturaleza química, de diversos tipos: aminoacídicas,
peptídicas,
proteínicas, glucoproteínicas y esteroideas. Esta diferente
composición química
explica que para la síntesis de cada hormona existe un tipo
especializado de
célula endocrina, de ahí la relación que se observa entre el
número de células y
hormonas en las diferentes glándulas endocrinas.
POR EL MODO DE SEGREGAR.
En algunas glándulas endocrinas se produce la secreción
inmediata de la
hormona, casi con la misma velocidad que se sintetiza, tal
como ocurre en la
corteza suprarrenal.
En otras las células acumulan la secreción en gránulos, que
se liberan cuando es necesaria.
Y en otros casos el acúmulo extracelular de la secreción da
lugar a la
formación de los folículos, cuya secreción se almacena en la
cavidad de esta
estructura y se libera cuando es necesario, un ejemplo
típico es la glándula
tiroides.
ORGANIZACIÓN HISTOLÓGICA DE LAS GLÁNDULAS INDEPENDIENTES
COMO ÓRGANOS MACIZOS.
GLÁNDULAS EXOCRINAS.
Las glándulas exocrinas independientes que conforman órganos
macizos,
tienen dos porciones fundamentales: el parénquima y el
estroma. El
parénquima es la parte funcional que caracteriza la glándula
y está constituido
por los elementos que derivan del epitelio glandular: las
unidades secretoras y
los conductos excretores; mientras que el estroma es la
parte que brinda
soporte y sostén y está constituida por el tejido conjuntivo
organizado en forma
de cápsula, tabiques, hilio y red reticular.
Las glándulas exocrinas se hallan rodeadas por una
condensación de tejido
fibroconjuntivo, la cápsula, de la cual se extienden
tabiques de tejido conjuntivo
hacia el interior de la glándula, que es por donde
transcurren vasos sanguíneos
y linfáticos, los conductos y los nervios. Estos tabiques de
tejido conjuntivo más
laxo, dividen a la glándula en lóbulos, los que a su vez se
subdividen en
tabiques más finos para dar lugar a los lobulillos. El hilio
es un engrosamiento
de tejido conjuntivo por donde penetran y salen de la
glándula los mismos
componentes que transcurren a través de los tabiques. La red
reticular de
fibras de tejido conjuntivo brinda soporte y sostén
directamente a los elementos
que constituyen el parénquima, a los vasos sanguíneos,
linfáticos y nervios
situados en el interior de los lobulillos.
Los vasos sanguíneos y linfáticos y los nervios,
generalmente siguen la
distribución del tejido conjuntivo. Estos capilares forman
redes alrededor de las
células secretoras y de los conductos terminales. Las fibras
nerviosas
terminales se ramifican y sus ramas finales terminan en
pequeños
engrosamientos sobre la superficie de las células secretoras.
El sistema de conductos de las glándulas exocrinas comprende
el conducto
principal, el cual se divide formando los conductos
interlobulares, los que a su
vez se ramifican y dan origen a los conductos
interlobulillares; estos se
continúan con los conductos intercalares, que son los más
pequeños y
constituyen las porciones de dicho sistema adyacentes a las
unidades
secretoras o acinos de la glándula.
El epitelio que reviste al sistema de conductos varia desde
el tipo simple plano
o cúbico en el conducto intercalar, aumentando el grosor del
epitelio al cúbico y
cilíndrico, al estratificado cúbico o cilíndrico en el
conducto principal. A medida
que el conducto disminuye de calibre los elementos de sostén
son más finos (red de fibras reticulares) y los conductos mayores están
rodeados por tejido
fibroconjuntivo con un revestimiento de musculatura lisa.
La secreción exocrina está controlada por el sistema
vegetativo y, en algunos
casos, ciertas hormonas actúan estimulando la secreción de
las glándulas.
GLÁNDULAS ENDOCRINAS.
Las glándulas endocrinas independientes, consideradas como
órganos
macizos, también presentan la organización histológica de
estroma y
parénquima. El estroma, igualmente está constituido por
cápsula, tabiques y
red reticular, pero el hilio no es evidente. El parénquima
solo está constituido
por la porción secretora, ya que carecen de conductos
excretores.
Las glándulas endocrinas se caracterizan por haber perdido
su unión con el
epitelio que las originó, por lo cual, están desprovistas de
conductos excretores
y la secreción la vierten directamente a la corriente
sanguínea o linfática.
Suelen estar constituidas por grupos de células que se
disponen en forma de
acúmulos, cordones y folículos, incluidos en un tejido de
sostén integrado por
fibras reticulares finas y asociados con una red sinusoidal
o capilar.
Las glándulas endocrinas están reguladas por el sistema
nervioso, o bien por
otras glándulas endocrinas o por combinación de factores
nerviosos y
endocrinos.
Glándulas mixtas Las glándulas se clasifican como mixtas
cuando su epitelio
glandular es exocrino y endocrino. En este tipo de glándulas
ambas funciones
pueden ser cumplidas, ya sea por un solo tipo de célula o
por diferentes
células.
En el hígado, las células hepáticas o hepatocitos segregan
simultáneamente
bilis en el sistema de conductos hacia el exterior y otras
sustancias
directamente hacia los capilares sinusoidales.
En el páncreas, células diferentes realizan cada función:
las exocrinas forman
acinos, los cuales son drenados por conductos, mientras que
la porción
endocrina constituye los islotes pancreáticos (islotes de
Langerhans) que,
segregan los productos hormonales (insulina y glucagón)
hacia los capilares.
De manera análoga actúan los testículos y los ovarios, un
grupo de células son
segregadas hacia el sistema de conducto (espermatozoides y
óvulos), mientras
que la secreción de tipo hormonal endocrina (testosterona y
progesterona)
respectivamente, pasa de forma directa a los capilares.
CORRELACIÓN HISTOFISIOLÓGICA EN EL TEJIDO EPITELIAL.
El tejido epitelial, como hemos estudiado se diferencia en
epitelios de cubierta y
revestimiento y en epitelio glandular.
Las membranas de cubierta y revestimiento por la organización
que tienen sus células son tejidos limitantes, ya que pueden formar verdaderas
barreras
celulares.
Las variaciones en el número de capas, formas celulares,
especializaciones y
otras estructuras presentes en los diferentes tipos de
epitelios, se
corresponden con los requerimientos funcionales y con una
amplia gama de
fuerzas físicas y químicas, a las que están sometidas las
superficies epiteliales.
Sus funciones principales son protección, absorción,
secreción e intercambio.
Para cumplimentar la función protectora, se requiere un
epitelio que presente
varias capas, o sea, un epitelio de tipo estratificado, ya
que esos son epitelios
difíciles de atravesar por gérmenes patógenos y también son
más resistentes al
desgaste. Así veremos que la piel, por ser un órgano tan
expuesto al medio
externo, va a presentar un epitelio estratificado plano
queratinizado. Sin
embargo, los epitelios simples cumplimentan funciones muy
diferentes.
Los epitelios adaptados para el intercambio, tanto de
líquido como de gases,
son epitelios simples planos, cuyas células presentan poco
citoplasma: por
ejemplo, el epitelio simple plano de los alvéolos
pulmonares, donde se lleva a
cabo un rápido intercambio de O2 y CO2, y el epitelio simple
plano del asa de
Henle en el riñón, donde se efectúa la reabsorción de
líquido del filtrado.
La absorción, por su parte, trae como resultado el
desarrollo de
microvellosidades, que aumentan la superficie de la membrana
plasmática y la
altura de la célula, por lo cual, los epitelios absortivos
como el del intestino
delgado, son de tipo cilíndrico.
Por último, el epitelio de transición, dadas sus
características morfológicas,
está adaptado para resistir la distensión, la hipertonicidad
y la especial
composición de la orina.
La función secretora implica un mayor desarrollo de los
organitos
citoplasmáticos, lo cual se corresponde con un incremento en
la masa
protoplasmática de las células.
Las células secretoras de proteína, por ejemplo, presentan
abundante RER,
aparato de Golgi y gránulos secretores; son células cúbicas
y en ocasiones
cilíndricas.
Los tejidos epiteliales se relacionan íntimamente con el
tejido conjuntivo, del
cual dependen para el mantenimiento de sus funciones. De él
reciben soporte,
sostén, nutrición, irrigación, drenaje y defensa y le
aportan protección. En
general se observa entre ambos tejidos la membrana basal,
que contribuye a
su unión entre ambos, y actúa como una barrera de
intercambio selectivo. Las
membranas epiteliales y glándulas independientes descansan
sobre una capa
de tejido conjuntivo vascularizado que recibe el nombre de
lámina propia o
corion, que junto con el tejido epitelial forma las mucosas
en los órganos
tubulares, que se relacionan con el exterior.
Los epitelios en general no presentan vasos sanguíneos,
nutriéndose por la difusión de las sustancias nutritivas provenientes del
tejido conjuntivo que
atraviesan la membrana basal.
La inervación de los epitelios es mediante terminaciones
nerviosas libres que
forman una red intraepitelial.
Las células epiteliales tienen vida limitada y se renuevan
constantemente como
resultado de una actividad mitótica continua. La velocidad
de renovación varía
desde 2 a 5 días en el intestino hasta más de 50 días en las
glándulas
salivales.
Las células epiteliales en determinadas condiciones
patológicas pueden sufrir
alteraciones reversibles y dar origen a un nuevo tipo de
epitelio, este fenómeno
recibe el nombre de metaplasia. Por ejemplo en los fumadores
crónicos la
acción irritante del humo produce la sustitución del
epitelio seudoestratificado
de la tráquea y de los bronquios por epitelio estratificado
plano. La falta de
vitamina A también produce la sustitución del epitelio de
los bronquios o vejiga
por plano estratificado queratinizado.
El control de la función glandular puede ser intrínseco y
extrínseco. El control
intrínseco de tipo celular es por un mecanismo genético que
permite la
producción de determinada secreción. El control extrínseco
puede ser nervioso
y hormonal. Ambos se realizan mediante sustancias
denominadas mediadores
químicos que a su vez se clasifican en neurotransmisores si
son producidos por
células nerviosas y hormonas si son producidos por células
glandulares. Estos
reaccionan con receptores intracelulares y de membrana que
estimulan directa
o indirectamente, respectivamente, los genes
responsabilizados con la
secreción.
CÉLULAS EPITELIALES ESPECIALIZADAS.
El proceso de diferenciación y especialización en el tejido
epitelial determina
que sus células adquieran ciertas características
estructurales y funcionales, en
correspondencia con la división del trabajo tisular, que se
produce, como
expresión de la dependencia y complementación celular.
Las principales células epiteliales especializadas son las
que realizan el
transporte activo de iones, como las células de los túbulos
renales; las que
transportan por pinocitosis moléculas a través de las
membranas tal y como
ocurre en los epitelios y mesotelios; los que secretan
proteínas, similares a las
células de las unidades secretoras serosas del páncreas y
glándulas salivales;
las que secretan polipéptidos de naturaleza hormonal, como
las células del
sistema APUD que intervienen en la captación de precursores
aminados y en
los procesos de descarboxilación en diversos órganos; los
que secretan mucus,
como las células caliciformes; las que secretan esteroides
de naturaleza
hormonal, que se producen en varios órganos como los
testículos, ovarios y
suprarrenales; y las mioepiteliales, presentes en las
glándulas sudoríparas,
mamarias y lagrimales alrededor de las unidades secretoras y
conductos
pequeños, donde por su contracción favorecen la expulsión de
la secreción. CÉLULA ABSORTIVA.
Por su importancia estudiaremos un modelo de célula
absortiva, especializada
en el transporte por pinocitosis y en el transporte activo
de iones, que integra
los elementos esenciales propios de las células absortivas
del epitelio intestinal
y de los túbulos proximales del riñón.
Las células absortivas en general son altas de aspecto
columnar o piramidal
con núcleos ovalados o esféricos situados hacia la base.
El proceso de diferenciación y especialización de estas
células determina
cambios estructurales en relación con la membrana celular y
los organitos
citoplasmáticos, en correspondencia con la función que
realizan.
Estas células presentan tres superficies: libre o apical,
lateral o intercelular y
basal, observándose en cada una diferentes especializaciones
de la membrana
celular.
En la superficie libre o apical presentan microvellosidades,
vistas al M/E, que
se observan como una chapa estriada al M/O, cuyo borde es
PAS positivo
producto del glicocalix asociado a las microvellosidades.
Esta cubierta
glicoproteíca formada por las propias células contiene
numerosas enzimas que
intervienen en la degradación de los compuestos que deben
absorberse. Se
observa además, una red de finos filamentos en el centro y
en la base de las
microvellosidades, constituido por filamentos de actina y
miosina que producen
el movimiento de contracción y acortamiento de las mismas.
El acúmulo de
filamentos en la porción apical de la célula, inmediatamente
por debajo de las
microvellosidades, recibe el nombre de velo terminal. Debajo
de las
microvellosidades también se observan numerosas vesículas
pinocíticas,
expresión de la intensa absorción que existe. Las
microvellosidades
incrementan la superficie disponible y los procesos que
facilitan la absorción de
macromoléculas.
En la superficie lateral las especializaciones presentes son
los medios de unión
intercelular. En el extremo apical de la superficie lateral,
próxima al borde libre,
se observa al M/O la barra terminal, que al M/E se denomina
complejo de
unión. Según varios autores, el complejo de unión está
constituido por tres
medios de unión: a) unión íntima o zónula ocluyente, b)
unión intermedia o
zónula adherente y c) desmosomas o mácula adherente. Más
hacia abajo se
encuentran las interdigitaciones y otros desmosomas
aislados.
En general todas estas especializaciones contribuyen a
mantener unidas
firmemente las células epiteliales entre sí, sobre todo en
la porción próxima a la
superficie apical.
En la superficie basal se observan las invaginaciones
basales de la membrana
celular, entre las cuales se orientan longitudinalmente las
mitocondrias. Estas
invaginaciones aumentan la superficie de intercambio activo
de sustancias e
iones, lo que a su vez explica, por la energía que se
necesita, la abundancia de mitocondrias en esta zona. Además se encuentran los
hemidesmosomas, que
contribuyen a la unión de las células epiteliales con el
tejido conjuntivo
subyacente, a través de la membrana basal.
En relación con los organitos citoplasmáticos, se observan
ribosomas libres
dispersos entre el retículo endoplásmico, que forma una red
continua de
canalículos y sáculos. El RER es el más abundante, aunque
predomina en la
porción apical de la célula el REL. El complejo de Golgi
está bien desarrollado
en posición supranuclear y los ribosomas también están
presentes,
particularmente en las células más viejas. Las mitocondrias
son alargadas,
numerosas y orientadas longitudinalmente, sobre todo en la
porción basal de la
célula.
Las moléculas de carbohidratos y aminoácidos se absorben,
desde la luz del
órgano y pasan a través del citoplasma hasta los capilares
sanguíneos
contenidos en la lámina propia. Las moléculas de lípidos
(ácidos grasos y
monoglicéridos) son absorbidas y procesadas por el REL
sintetizando
triglicéridos y por el Golgi que le incorpora el componente
proteico. Las goticas
de lipoproteínas así formadas (quilomicrones) pasan entonces
lateralmente al
espacio intercelular, desde donde descienden, atraviesan la
membrana basal y
penetran en los capilares linfáticos.
El transporte de agua es muy discutido pero, en general, se
acepta que es un
proceso osmótico secundario al gradiente de concentración
resultante del
transporte de iones, que se absorben en la superficie libre,
de los cuales
algunos pasan a través del citoplasma y otros siguen el
camino del espacio
intercelular. De forma general, la absorción y transporte
del agua e iones se
produce conjuntamente para mantener así un equilibrio
osmótico.
CÉLULA SECRETORA DE PROTEÍNAS POLARIZADA.
El proceso de diferenciación en las células especializadas
en la secreción de
proteínas determina cambios estructurales en correspondencia
con esta
función. Por su importancia estudiaremos un modelo de célula
secretora de
proteínas, que analiza los componentes esenciales propios de
este tipo de
células que integran las unidades secretoras serosas del
páncreas y las
glándulas salivales.
Estas células, en general, son piramidales o poliédricas con
un núcleo esférico
situado ligeramente hacia la base. Presentan abundante RER y
mitocondrias
situadas debajo y a lados del núcleo. Por la abundancia de
RER, al M/O, la
porción basal de la célula presenta una intensa basofilia.
El complejo de Golgi
está muy desarrollado y se observa en posición supranuclear.
Son evidentes
los tres componentes del Golgi: vesículas de transferencias,
sacos aplanados y
vacuolas de condensación. En la porción más apical de la
célula son muy
abundantes los gránulos de secreción, que por su composición
se denominan
gránulos de cimógeno. Los aminoácidos provenientes de los
capilares
sanguíneos pasan al interior de la célula, proceso que es
acelerado por una
bomba de transporte activo. Una vez en el citoplasma, a
partir de los mismos,
los ribosomas adheridos al RER sintetizan proteína de
secreción que pasa, por medio de las vesículas de transferencias a los sacos
aplanados de la cara
formadora del complejo de Golgi, donde comienza a
condensarse la secreción.
Mediante un proceso continuo de desplazamiento ascendente de
los sacos, en
el extremo secretor de dicho complejo, se forman las
vacuolas de
condensación, que dan lugar a los gránulos de secreción.
Estos gránulos se acumulan en la porción apical tornándose
cada vez más
densos debido a la pérdida de agua. Son los responsables de
la acidofilia que
se observa al M/O en la zona apical de la célula.
Posteriormente estos gránulos
se liberan por un proceso de exocitosis a través de la
membrana en el polo
secretor de la célula.
Si en lugar de proteínas la secreción es una glucoproteína
el componente
carbohidratos puede ser añadido a nivel del RER o del Golgi.
La energía necesaria para todo este proceso de secreción es
aportada por las
mitocondrias.
En el capítulo, que aborda en detalles la célula, puede
profundizar en la
estructura y la función de las especializaciones de la
membrana y de los
organitos citoplasmáticos que hemos considerado, de manera
general, en los
modelos de célula absortiva y secretora de proteínas
estudiadas como células
especializada.

Glándula exocrina
|
Sistema exocrino
|
|
|
|
|
Estructuras
básicas
|
Glándula exocrina.
|
|
Glándula exocrina
|
|
|
|
|
Glandula exocrina
|
|
|
|
|
|
|
|
Sinónimos
|
|
Glándula de secreción externa.
|
|
Enlaces externos
|
|
|
|
Las glándulas
exocrinas son un conjunto de glándulas que
se distribuyen por todo el organismo, formando parte de distintos órganos y aparatos que producen diferentes sustancias
no hormonales que
realizan una función específica, como las enzimas. Las
glándulas exocrinas también se llaman glándulas de secreción externa.
Las glándulas
exocrinas secretan productos químicos a través de conductos o tubos que llevan
las secreciones a las cavidades corporales, a la luz de un órgano o a la
superficie corporal, a diferencia de las glándulas endocrinas que llevan su producto
hacia el líquido intersticial circundante más que
hacia conductos. En algunas glándulas exocrinas se puede distinguir una parte
productora o secretora de la sustancia y otra parte excretora o que vehiculiza
la sustancia a un lugar determinado.
Los tejidos que
producen hormonas se
pueden clasificar en tres grupos: Glándulas endocrinas, cuya función es la
producción exclusiva de hormonas; glándulas exocrinas, que producen
también otro tipo de secreciones además de hormonas; y ciertos tejidos no glandulares,
como el tejido nervioso del sistema nervioso autónomo, que produce sustancias
parecidas a las hormonas.
El sistema
exocrino es el conjunto de glándulas exocrinas que están distribuidas
por todo el cuerpo y que, generalmente, no tienen conexión ni función en común
entre ellas.
Índice
- Glándulas unicelulares: están formadas por una sola célula secretora
como las células caliciformes o mucosas que se
encuentran distribuidas entre las células cilíndricas del epitelio de
muchas mucosas como
la del estómago.
- Glándulas pluricelulares: están formadas por múltiples
células, formando estructuras más o menos complejas, adoptando morfologías
características como:
- Túbulos o glándulas tubulares: la
parte secretora tiene forma de tubo.
- Alveolos o glándulas alveolares: la
parte secretora tiene forma de bolsa o alvéolo.
- Acinos o glándulas acinosas: la
parte secretora es un conjunto de bolsas que drenan en uno o varios
túbulos.
- Mixtas: es la combinación de las
anteriores: tubuloalveolar, tubuloacinar, etc.
Según la
estructura
- Tubular (única recta) simple.
- Tubular (arrollado) simple.
- Tubular simple contorneada.
- Alveolar (única) simple.
- Alveolar (múltiple) simple.
- Alveolar múltiple compuesta.
- Tubular múltiple compuesta.
- Tuboalveolar compuesta.
- Glándula simple: si el conducto excretor
es único.
- Glándula compuesta: si el conducto
excretor está ramificado.
Según el
producto de secreción[editar]
- Glándulas mucosas: corresponde a una secreción rica
en carbohidratos, llamada mucina. Tiene
una consistencia viscosa con función lubricante o protectora.
- Glándulas serosas: producen una secreción serosa,
que es acuosa, fluida y rica en proteínas de
naturaleza enzimática.
- Glándulas seromucosas: producen secreciones mixtas,
con viscosidad intermedia.
Muchas de las
proteínas secretadas por las glándulas exocrinas son enzimas.